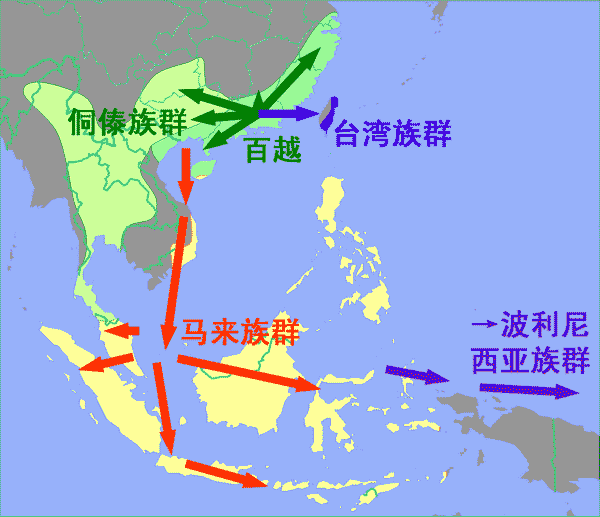
�ϵ�������Ⱥ��׳����Ⱥ��ϵ�Ľ����Ŵ���ϵ
��ԡ��IJ�������پ���ޱ������żѹ�-����������������졢��������������������������꿡���ͤƽ��¬���塢��ľ��������-������������-ɣ����̸���塢����
ժ Ҫ (�й�������.��������ѧ��̳.baiyueren����)
�������ϵ������ڶ����ǡ������ݺ�ӡ���������չ������ϵ�����������Ե�������Ϊ�����������ϵ����������-���������Ǻ�̨������������ϵ������Ի��������������̨�����ϵ���ԭ��ļ�˵��������ѧ�ҡ�����ѧ�ҡ�����ѧ�Һ��Ŵ�ѧ���ж������ۡ��ձ鹫�϶����ϵ����ˣ��ܿ����������˺Ͳ����������ˣ�Դ�������ϵ����ˣ������ǵ����̨��ԭס�����Ҵ�½�ϵ�׳����Ⱥ�ٶ�Ϊ�����ϵ����˵�Դͷ��
������ڱ����У������о���30���й�׳����Ⱥ��23��ӡ�������Ǻ�Խ�ϵ�����-������������Ⱥ����1509��YȾɫ��ķ���������20��SNPλ���7��STRλ�㡣������Ⱥ���ڸ�ϵ�Ŵ���ϵ������ʾ�����֮ͬ������Ϸ���֤ʵ��׳����Ⱥ���Ŵ����漸��δ���й�����Ӱ�죬���ҹ�����ӡ���˵��Ŵ������塣O1a*�ڴ���Ⱥ�������и߷�������������ϵ��Ⱥ��ʮ���ټ���O1a*����Ⱥ������ͼ������ӡ���˵���ϵ����������ѧ�о����������Դ��̨��ԭס��������׳����Ⱥ��
���ۣ����Ƿ��֣���̨��ԭ���˵�෴�������ǵ�����Ⱥ�ڸ�ϵ�Ŵ���ϵ�ϲ���̨����Դ�����⣬���ǵĻ�����̨��ԭס���ӡ�����ڸ�ϵ�Ϻܿ��ܶ�Դ��׳����Ⱥ������֧��Ⱥ�ƺ��������˶�����չ�Ĺ��̣����ǵ��о������ʾ�Ŵ���֧��һ������̨��ԭס��׳����Ⱥ������--������������Ⱥ���ڵĴ���Ⱥ��
���
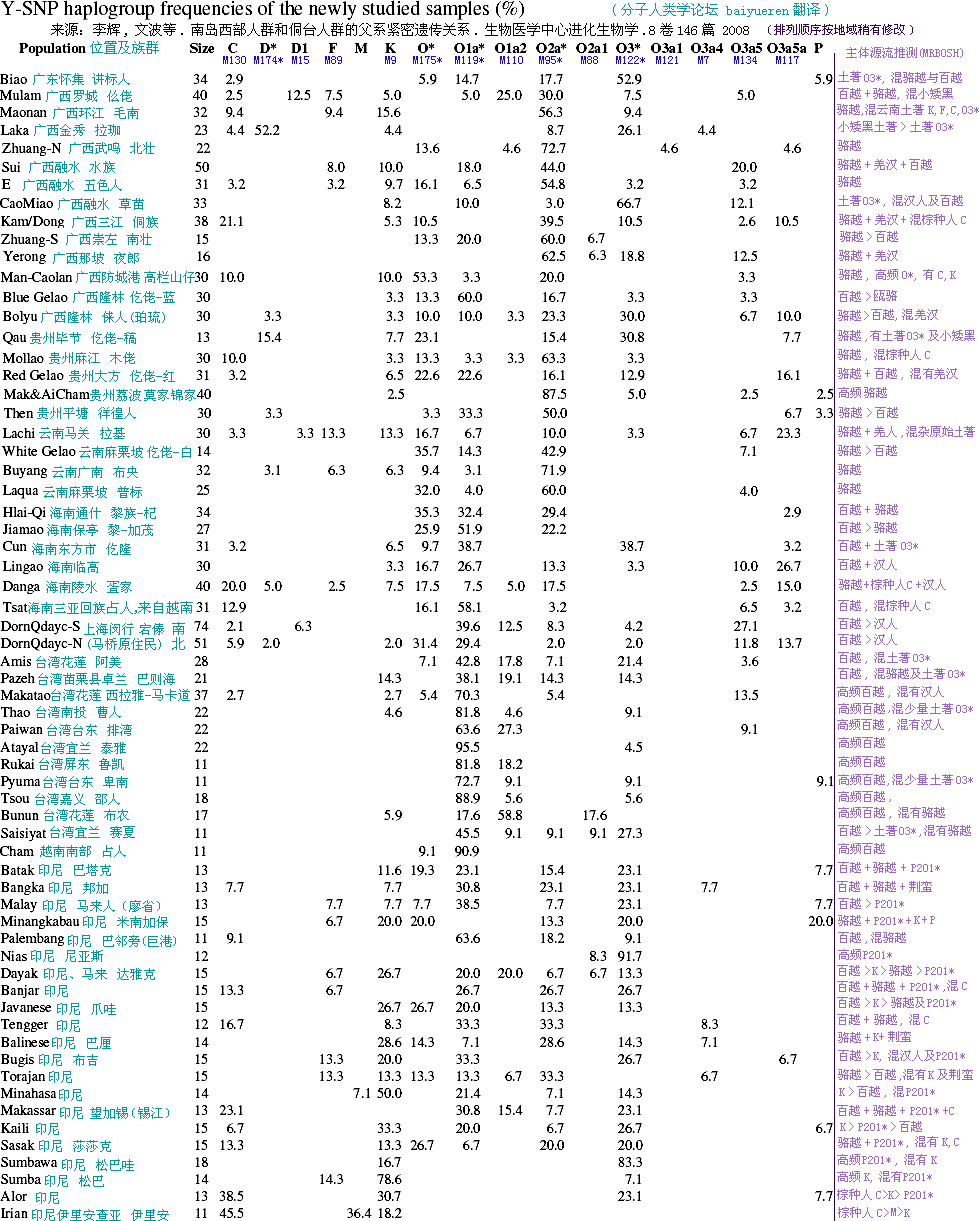
Ϊȷ��׳����Ⱥ���ϵ�������Ⱥ���Ŵ���ϵ�����Dzⶨ��23�������ǵ�����Ⱥ��11��̨��ԭס��Ⱥ��Ⱥ�����ڵؼ�ͼ1����Ⱥ��Ϣ����1����1509��YȾɫ��ķ���������20�����������̬��λ���7���̴��ظ�����λ�㡣�����о������˼���ȫ���й�׳����Ⱥ��̨��ԭס��Ⱥ��������
���⣬��������˶������ж��ǺͶ��������Լ������ڵ�134��������ȺSNPλ�㶨�嵥��Ⱥ�����ɷ֣�PC�������������ʾ���������ǺͶ�����Ⱥ����ȣ�׳����Ⱥ��Ϊ�ӽ������ϵ���Ⱥ��ͼ2��������׳����Ⱥ�������ϵ���Ⱥ�кܽ����Ŵ���ϵ��׳��-������-̨�弯�Ŵ�������Ⱥ�еķ��������ɷ�2�и߶�����ԣ��������1�������⣬O1a-M119��̨��ԭס������Ƶ���Ⱥ��ƽ��77���������ֵ��54����100�����ȣ�����2��O1a*��O1a2֮�ͣ���������������Ⱥ����Ȼ��<5��������ˣ�Ԥ��O1a-M119��Ϊ����׳����Ⱥ�������ϵ���Ⱥ�Ĺ�ϵ�ṩ�ḻ����Ϣ��

ͼ2�е����ɷ�ͼ��ʾһЩ׳����Ⱥ�ӽ�������Ⱥ�Ĵء���������Ϊ׳����Ⱥ�ͺ�����Ⱥ�й�ͬ�����ȣ����ܵ����������Ŵ���������֮����Ȼ�����Դ�Ҳ��������һ�ֽ��ͣ����Ƕ��Ǵ�½�ϵ�׳����Ⱥ���Ŵ��Ͽ����ܵ����й������Ӱ�죬��Ϊ���Ǵ�2500��ǰ�Ϳ�ʼ���ڶ��ӡ���Ϸ������������ٶ��ִ�׳����Ⱥ��׳���������ȵı�������һЩ�������׳����Ⱥ������Ϊ��Ϸ����еĸ�ϵ��Ⱥ�����ϵ�ԭס��Ⱥ�����塢��ï�˺���¡�ˣ���̨��ԭס������Լٶ�Ϊ��Թ����ģ���Ϊ���ǵ��Ļ������ܵ���½�ϵ������Ļ���Ӱ�졣���ԣ����Ŵ��ṹ����Щԭס������ܺ�������׳�����ȡ�
Ϊ�����ٶ��Ĵ�½׳����Ⱥ�еĺ����Ŵ�Ӱ�죬���ǽ���½׳����Ⱥ������ԭס��̨��ԭס����й������Y-SNP����Ӧ���ڻ�Ϸ����С�Ϊ�˷��������ǽ���3��Ⱥ���������Ϊ��½׳����Ⱥ�ĸ�ϵȺ�塣�����ʾ����ԭס����Ŵ����Ǹ�(2.145��0.927)��Ȼ���й�����(-0.314��0.422) ��̨��ԭס��(-0.831��0.662)�Ĺ�����û�С������ADMIX����������Ŵ�����ֵ��ʾ���ִ�׳����Ⱥû�й��Ŀ��ܡ���������ʾ׳����Ⱥ�ĸ�ϵ�Ŵ���ϵ���൱�����ģ�׳����Ⱥ�������ϵ���Ⱥ֮����Ŵ���ϵ����δ����Ⱥ���ӵ�Ӱ�졣
�����ǵ�����Ⱥ����Ҳ�ǻ�ϵġ������ǵ��о��У����趫���ǵ����ڸ�ϵ�Ŵ�����������Ⱥ��Ļ�ϣ�׳����Ⱥ��̨��ԭס���Լ��ܽӽ��Ͳ����˵�����Ⱥ�������������������ӡ���������˵Ļ�Ϸ��������ڷ����м����˴������еõ��İͲ�����������Ϊ��ϵ��Ⱥ�ṹ֮һ�����ǵķ�����ʾ���µĻ�ϱ�����׳��Ⱥ��(0.713��0.124)��̨��(0.143��0.125)�ͰͲ����� (0.144��0.050)������׳�����ȶ�ӡ���˵ĸ�ϵ�Ŵ���������ǿ�Ƶġ�����Щ�����д���һЩ��ȷ���ԣ���Ϊ���ǹ��ڶ����ǵ����ǻ�Ѫ�ļ��������õ�����ġ�
����O1a*��׳���������ϵ���Ⱥ�����ɫ�ĵ���Ⱥ������ʹ��O1a*�����7��STRλ��������׳����Ⱥ��ӡ���˺�̨��ԭס��֮����Ŵ����룬���Ŵ������ͼ3��STRԭʼ���ݼ�����2�������ǵ��о���ʾ̨���ӡ����Ⱥ֮���Ŵ����������ģ����������׳��Ⱥ���̨��ԭס���Ŵ������3����׳��Ⱥ���ӡ����֮����Ŵ�����ܽӽ�׳����Ⱥ��̨��ԭס��ľ��롣��Щ���ֱ���ӡ���˺�̨��ԭס�����Ŵ��϶���׳����Ⱥ�ĸ��ӽ��������������������ϵ�Ⱥ��֮��ľ��롣���⣬��O1a*�����7��STRλ���ϵĶ�̬����׳�������б���ӡ���˺�ԭס���и��߷���ͼ3������̬����ߵ���Ⱥδ����������ϵģ�Ҳ���������ܱ���Ⱥ��Ѫ�Ľ�������ǣ�׳������O1a*����Ⱥ�ϸߵĶ�̬��Ӧ������Ⱥ����������ɵģ���Ϊ���ֵ���Ⱥ���ܱ���Ⱥ�м��������ڣ����Բ��ǻ�Ѫ�����˸��ߵĶ�̬�ԡ�������ȡ�ö�̬�Ժͷ���ʱ������ʾ��׳����Ⱥ�ܿ�����ӡ���˺�̨��ԭס��ϵ�Ŵ���ϵ�ϵ�����Ⱥ�塣������Y����Ⱥ����O3-M122��O2a-M95��Ҳ������O1a*���Ƶ�ͼ�ף���ʾ׳����Ⱥ��ӡ���˺�̨���˵��Ŵ�����Ⱥ�������Ⱥ֮��ľ�����ӽ�����3������Ȥ����O2a�������ܽ�һ���ݵ���������Ⱥ����������о�������Ĺ۵㡣

������������ȺO1a*�����7��STRͻ��λ�㹹�����н����磨ͼ3�������綫���ǵ����̨��ԭ���˵��THH������ģ�Ҳ����˵�����ǵ��������Դ��̨��ԭס����ô�����еĶ�������ϵ��̨����ϵ��Ӧ���й����ڵ�ͣ����ߣ����ߡ���ͼ3�У�׳����ϵ����ɫ�ڵ㣩��������������ġ����еĶ�������ϵ����ɫ�ڵ㣩��̨��ԭס����ϵ����ɫ�ڵ㣩ֱ�ӻ��Ǽ�ӵع�����������׳����ϵ���෴�أ�û��̨����ϵ������һ���������������ӵ���������ϵ�Ľڵ㡣��Щ���ֱ��������ǵ���ֱ��Դ��̨��ԭס��Ӧ���Ƕ����ǵ����̨��ԭס����Զ�������Դ��׳��Ⱥ�塣
���ǽ�һ���۲���������Щ�붫������ϵ������׳����Ⱥ��ϵ����Ȥ���ǣ����붫����������׳�������Ͳ������Ժ��ϣ��������Ժ���������Ĺ�����ͼ3�д�������Ȧ����ɫ�ڵ㣩����Щ���Ϻ�����Ⱥ��λ�ڱ������ܱ����������壬Gulf of Tonkin�����ر���Խ���ϲ�����������-������������Ⱥ��ռ�ˣ�cham�����Լ����ϵĻػԣ�Tsat�������Կ�����������������ϵ��׳����Ⱥ��ӡ���ˡ��������Ǽ��趫���ǵ�����Ⱥ��Դ�ڱ������ܱߵ��������Ǵ���ӡ֧�뵺�������뵺����Ǩ�㣬��������ɢ��̫ƽ���ӡ����Ĵֵ��졣
���ǹ�����������O1a*����Ⱥ����������������33765��5221�꣬��ĩ�α������Ǻϡ�����������׳��ϵ�����������33193��5577�꣬��O1a*����������ܽӽ�������̨����׳���ĵ����ʹ�Χ���������ѹ���̨��ص���ʵ�������Щ����������������׳����Ⱥ��̨��ԭס��Ⱥ�Ķ���������ǹ�������������Ǹ�̨��ص���14659��3110�ꡣ����̨�����������������ֵΪ21268��3148�ꡣ��Ȥ���ǣ�����������ֵ�ܽӽ�̨���ѷ��ֵ�����������Ŵ棬�������ˡ��ɴ������ƶ�O1a*����Ӵ�½��̨�嵺����������ھ�ʯ��ʱ����
�����������п��Թ۲쵽����������ȷ�Ķ����ǵ��쵥���͵Ĵأ����Ǽ��������ߵ��������ߵĶ����Ǵص������9895��2393�꣬�����ұߵĴ�ȴ��25880��7137�ꡣ����ѧ�϶����������������������Դ����Ĺ��Ʊ����ǵĹ�ֵҪ����ö࣬��Լ5000-6000��ǰ������ɷ����������е�׳�������ͺͶ����ǵ��쵥������һЩ������˵���ڶ�����������׳������Ⱥ��������������п��ܷ�����ƿ��ЧӦ������ѧ�ϵ�ƿ��ЧӦ����������խԽ�Ϻ���������O1a*����Ⱥ�ܿ�����������������������7500�����֮ǰ�ɴ������ǵ�����ɢ����������ʱ��O1a���������ǵ�������Ŀ������Dz��ܺ��ӵģ���Ϊ�Ŵ�ѧʱ����Ƶľ��Ȳ������ų����ֿ����ԡ�
Ӧ��ע����ǣ��ڡ��ر�쳵����˵�д������ֲ�ͬ�ĽǶȣ���1���������Դ��Ҳ����̨��ԭ��˵���Լ���2������ģʽ��Ҳ���Ǵ�ӡ�Ὺʼ�Ŀ�����ɢ�����о��У�Ϊ��̨֤��ԭ��˵�����Ǽ������ǰ�о��к����漰�İ���׳�����˺Ͷ����ǵ������ڵ������ϵ���Ⱥ�����Ƿ���̨�岻���Ƕ�����ӡ���˵�ԭ�磬���ٲ����丸ϵ������ϵ�Ĺ��硣����̨��ԭס���ӡ���˶�Դ��׳����Ⱥ�������߸�ϵ������ϵ���ǵ�һ��Դ������ֻ�����������¼���
����˼���ǣ����ǵ��о����������ڶ�����Ⱥ����̫ƽ������ķֲ��������ϵ���Ⱥ�ֲ�����һ�¡�̨�弰Զ���ܿ��������ǵļ�����ֱ����Դ�ڶ��Ǵ�½������Щ������Ⱥ���Ͳ��������ǵļ���������ӡ֧�뵺����������DZ�����Ⱥ��������Ǩ��������ģ��Ǿ���ζ������ͬ������˳����������·�ߵִﶫ���Ǻ�̫ƽ���졣
��ʵ�����ǵļ�������̩��Ⱥ��һ��Դ������һ�£���̩��Ⱥ��׳����������������������̨����������ɡ����ǵļ����������������о�����׳����Ⱥ�Ͷ����ǵ�����Ⱥ���о��ϵ���Ⱥ����Դ�Ǿ��Ա�Ҫ�ġ������������ЩȺ�壬���������Ǻ�̨��ԭס����Ե��ʮ�ֽӽ����Ӷ����������ϵ��˶�����Դ��̨��Ľ��ۡ�
���ǵĽ�����������弫�п����Ƕ����ǵ�����Ⱥ��ԭ�硣��������Ⱥ���ӡ�ᶫ����̫ƽ����������̵ĸ����ԣ��Լ������Ͷ����ϵ���Ⱥ���Ŵ���������������������о���û��ѡ���������ǵ����ݡ���Ϊ���������ֻ�����������ϵ�Ⱥ�塣������������Ⱥȱ��O1a-M119����ʵ�����˺���ģ����Ҳ��ܼ���ƿ��ЧӦ������������������YȾɫ�嵥���ͱ��ֳ��߶�̬�Ե�ԭ��
�����ǶԸ�ϵ�Ŵ���ϵ�ķ�����һ�£��������뵺Ⱥ���������DNA�о�Ҳ��������Լ��ĩ�α��ڣ�Last Glacial Maximum��ʱԭʼ�����˵����Ⱦ�ס��ӡ֧�뵺��������֧����Ⱥ�徭�������뵺��ɢ�������ǵ��졣�Զ����ǵ�����Ⱥ��mtDNA�о�Ҳ��������ʹ������һ���ϵ����˴�̨����������˿�ͳ��ѧ��Ҳ��һ����Ҫ���¼���
���ǵĴֽ��۶��ǻ��ڶ�O1a*�ķ�����һ������Щ��ȺYȾɫ����ϵ��ֻռ�ٲ��ֵĵ���Ⱥ��O1a*��ϵ��Ƶ����̨����Ⱥ�������߷��������������������������˺�׳����Ⱥ�в����ر�ࡣһЩ��Ⱥ�¼����ܻ��漰��������YȾɫ����ϵ����������Ҫ�ġ���ͬ��Դ�ĸ�ϵ��ϵҲ������������˵�ϵ�Ⱥ�嵽��֮ǰ��ӡ��������Ⱥ�����и������������ǵ������ǺͶ�������֮���Ŵ���ϵ�ĸ��ӳ̶ȳ��������ǵ�Ԥ�ϡ�
����
���ǵĽ����ʾ���ڸ�ϵ�Ŵ���ϵ��׳����Ⱥ��������������Ⱥ����ӽ����������ϵ���Ⱥ��O1a-M119��׳����Ⱥ�������ϵ���Ⱥ����Ҫ��YȾɫ�嵥��Ⱥ����STR��̬�Ա��������ϵ�������Ⱥ�塪��̨��ԭס��Ͷ����ǵ������������Զ�������Դ��׳��Ⱥ�塣��ˣ������ǵ�����Ⱥ�ܿ�����Դ�ڱ������������׳�������ԭ�磬Ȼ��Խ������Ǩ����ӡ�������ǡ�����෴��̨��ԭס�����Ǵ��й���½ֱ���������ǵ��о�����������Ŵ��Ͽ����ƶϳ�һ������̨��ԭס��Ⱥ��׳����Ⱥ�����������������������ڵĴ�Ⱥ�塣
| ��Ⱥ | ������ | λ�� | ���� |
| Bolyu | �g��(����) | ����¡�� | �ϸ��������� |
| Yerong | ҹ�� | �������� | �����Bu-Rong |
| Qau | ����-�� | ���ݱϽ� | �����Ge-Chi |
| BlueGelao | ����-�� | ����¡�� | �����Ge-Chi |
| Lachi | ���� | �������� | �����Ge-Chi |
| Mollao | ľ�� | �����齭 | �����Ge-Chi |
| RedGelao | ����-�� | ���ݴ� | �����Ge-Chi |
| WhiteGelao | ����-�� | ���������� | �����Ge-Chi |
| Hlai-Qi | ����-� | ����ͨʲ | ��������� |
| Jiamao | ��-��ï | ���ϱ�ͤ | ���� |
| Buyang | ���� | ���Ϲ��� | �����yang-biao |
| Cun | ��¡ | ���϶����� | �����yang-biao |
| Laqua | �ձ� | ���������� | �����yang-biao |
| Man-Caolan | ����ɽ�� | �������Ǹ� | Be�CTai |
| Zhuang-N | ��׳ | �������� | Be�CTai |
| Zhuang-S | ��׳ | �������� | Be�CTai |
| Lingao | �ٸ� | �����ٸ� | Be�CTai |
| E | ��ɫ�� | ������ˮ | ��ˮ�� |
| AiCham | ���� | ������ | ��ˮ�� |
| Kam/Dong | ���� | �������� | ��ˮ�� |
| Sui | ˮ�� | ������ˮ | ��ˮ�� |
| Mak | Ī�� | ������ | ��ˮ�� |
| Mulam | ���� | ������ | ��ˮ�� |
| Maonan | ë�� | �������� | ��ˮ�� |
| Biao | ������ | �㶫���� | ��ˮ�� |
| Then | ������ | ����ƽ�� | ��ˮ�� |
| Danga | �D�� | ������ˮ | �� |
| DornQdayc | 崴�������ԭס�� | �Ϻ����� | �� |
| CaoMiao | ���� | ������ˮ | ��ˮ�� |
| Laka | ���� | �������� | ��ˮ�� |
| Amis | ���� | ̨�廨�� | �ϵ��������� |
| Pazeh | ���� | ̨���������� | �ϵ��������� |
| Siraiya-Makatao | �����š���������ƽ�ң� | ̨�廨�� | �ϵ��������� |
| Thao | ���� | ̨����Ͷ | �ϵ��������� |
| Paiwan | ���� | ̨��̨�� | �ϵ��������� |
| Atayal | ̩�� | ̨������ | �ϵ���̩���� |
| Rukai | ³�� | ̨������ | �ϵ��������� |
| Pyuma | ���� | ̨��̨�� | �ϵ��������� |
| Tsou | ���� | ̨����� | �ϵ������� |
| Bunun | ��ũ | ̨�廨�� | �ϵ��������� |
| Saisiyat | ���� | ̨������ | �ϵ��������� |
| Batak | ������ | ӡ�� | �����ϵ��� |
| Bangka | ��� | ӡ�� | �����ϵ��� |
| Malay(Riau) | �����ˣ���ʡ�� | ӡ�� | �����ϵ��� |
| Minangkabau | ���ϼӱ� | ӡ�� | �����ϵ��� |
| Palembang | �����ԣ��ۣ� | ӡ�� | �����ϵ��� |
| Nias | ����˹ | ӡ�� | �����ϵ��� |
| Dayak | ���ſ� | ӡ�ᡢ���� | �����ϵ��� |
| Banjar | �� | ӡ�� | �����ϵ��� |
| Javanese | צ�� | ӡ�� | �����ϵ��� |
| Tengger | �� | ӡ�� | �����ϵ��� |
| Balinese | ���� | ӡ�� | �����ϵ��� |
| Bugis | ���� | ӡ�� | �����ϵ��� |
| Torajan | �� | ӡ�� | �����ϵ��� |
| Minahasa | �� | ӡ�� | �����ϵ��� |
| Makassar | �������������� | ӡ�� | �����ϵ��� |
| Kaili | �� | ӡ�� | �����ϵ��� |
| Sasak | ɯɯ�� | ӡ�� | �����ϵ��� |
| Sumbawa | �ɰ��� | ӡ�� | �����ϵ��� |
| Sumba | �ɰ� | ӡ�� | �в��ϵ��� |
| Alor | �� | ӡ�� | �в��ϵ��� |
| Irian | ���ﰲ | ӡ�����ﰲ���� | ��������� |
| Cham | ռ�� | Խ���ϲ� | �����ϵ���ռ�� |
| Tsat | �ػԣ����壩 | �������� | �����ϵ���ռ�� |
����ѧ�����Ŀ�-һ�ߡ�3��2168ƪ �����ڶ�����ڴ��ĺ���ԭס��ϵ�Ŵ��ṹ
�����ڶ�����ڵĺ���ԭס��ϵ�Ŵ��ṹ
���,���,����Ө,½��,��Ԫ��,�,����,�����,��ʿ��,����
��ѡ���� (�й�������.��������ѧ��̳.baiyueren����)
Ҫ�㣺Ϊ���������ĺ��ϵ��Ŵ��ṹ���Ϸ���ڴ�������Ŵ��ṹ�������о���ȫ��6����Ⱥ��405������ԭס��YȾɫ�壬�����ܵ��½���½��Ⱥ�䶯�ͻ��ӵ�Ӱ���С���������DZ����ں���ԭס����O1a*��O2a*����Ⱥ��ռ���Ƶġ����⣬��½��ռ���Ƶĵ���ȺO3��ԭס���е�Ƶ�ʺܵͣ�˵�������ǹ�����Ⱥ�塣��������Ա�������ԭס����2����ǰ�����룬��������Ҫ�ĵ���Ⱥ���붫��֮��3.1-3.6����ǰ����

���������
�����ǵ��о��У����Ƿ����˺���ԭס��6����Ⱥ��YȾɫ����졣���Ǽ����405���������Ը����22�������ն�̬�ԣ�SNPs������7���̴��ظ���STR����̬�ԣ���ȷ���˻���YCC��ISOGG���������ĵ���Ⱥ�����ǽ�������Y��SNP����ȺƵ�ʷ���dz����Ƶ���Ⱥȡ����O1��O2���ڸ���Ⱥ�г������ģ����Һܿ����Ǻ���ԭס���ԭʼ����Ⱥ���ں���ԭס��֮һ������У���Щ����Ⱥ����Ƶ�ʴﵽ��100������������һ������ƿ��ЧӦ����Ⱥ����ʵ�ϣ����ֲ���ƫƧ��ɽ������������һ����С����Ⱥ��չ�����ģ����Y-STR��̬�Խϵ�֧��������۵㡣O1��O2����̨��ԭס��Ͷ������ϲ������ĵ���Ⱥ�����й��������ϲ��غ��������ݺͷ��ɱ����ϵ�����Ⱥ��O1��O2Ҳ�߷������ǵ�����������ѧ�Ͻ��Ƶ�׳������ϵ�����ɡ���׳����Ⱥ��ȣ�������Ⱥ��O1������Խϵͣ��ر�����ӡ֧�뵺�����IJ��������������Ⱥ�С�O3�Ǻ�������Ⱥ�ҵ�IDͬ�ĵ���Ⱥ�����壨50.51������������Ⱥ��54.70%�����ں���ԭס�����Ǻ��ٵģ�6.91��������Ȼ��̨��ԭס���е�Ƶ�ʴ�11.36����0��-37.6�������Լ��ڴ�½�Ϸ���׳��19.6�������в����ϸ���54.02����ԭס��Ⱥ���нϸߵ�Ƶ�ʡ��������̨��ʹ�½�Ϸ�������Ⱥ��Ⱥ���Ⱥ���뺺��������Ŵ����ӽ��١��������½��Ⱥ��ͬ��D��P��N��Q�ȵ���Ⱥδ�����ں���ԭס���С�
׳�������������Ⱥ��ɢ���ڴ�½�Ϸ��������ֲ������Ǽٶ�������֮���Ȼ�г�ֵĻ������������ҿ��Ա�����һ��Ⱥ�壬�������ǵ�Y-SNPƵ�ʵ����������Ƶġ�������Ⱥ�������ϲ�Ⱥ����ⲿ��֧���������е�ƿ���ͱ������������裬���Ǵ��������֧����ࡣ��������ʾ������ȫ��ͬ�ڴ�½�ϻ��ӵ���Ⱥ���ڱ���Ⱥ���У��ϸ��ޡ��й�����Ͳ���ϵ��Ⱥ�֮������ӽ��ģ���Ȼ���Ŵ����ϵ���Ͱ���̩��Ⱥ��ȫ��Ӱ�춼���Ժ�����ϵȺ�塣�����ҵ������Ⱥ���е��ⲿ��֧��̨��ԭס��ͺ�����¡�ˣ����ֳ���Խϵ͵ĵ���Ⱥ���졣���ǻ������Ϊʲô������¡�˺�̨��ԭס��ӽ���

���ɷַ���Ҳ��ʾ�Ϸ��ͱ�������Ⱥ�Ƿֿ��ġ������ɷ�ͼ�Ļ����ϣ�̨��ԭס��ӽ�������Ⱥ��������ԭס���Ƿ�Ϊ��֧��ͬ��Ⱥ�塣��¡�˿�����̨��ԭס��һ�𱻷ŵ�������Ⱥ�С����еĺ�������Ⱥ��dz��ӽ��ϱ��ֲ����ϲ���Ե���ٴ���֤�����ǹ������Ŵ��ṹ�����ַֻ�����ȷ����ʾ������Ǵ����ϲ���Ե��λ�á�
����SNP����������ԭס���뱱����Ⱥ�Ǹ���ģ��ر����ڻ�����Ⱥ���Ϸ�����ʱЯ���Ÿ�Ƶ��O3����Ⱥ���ϸ�����Ⱥ���ϵ��������ҲΪO3�ڻ��ϵ�Ƶ�ʵ����������˹��ס�����������о���ʾ�ϸ��ͺ�������Ⱥ�����Ĺ�ͬ�������й����ϲ����������������ᵽ�ģ�����ԭס����ȱ��O3�����Ǻ���ԭס��ռ���Ƶĵ���ȺO1��O2��ͬ��Ҳ�ڴ�½׳����Ⱥ��̨��ԭס����ռ���ơ�����ȺSNP�������ų⺣��ԭס���׳��-̨����Ⱥ֮��Ļ���������

����������У��漰������������̨��ԭס���Լ����ϱ��������ʡ�ݹ㶫��������׳������Ⱥ����ͼ3�����ÿ�����ˣ�����ԭס����ʾΪ��ɫ�ڵ㣩��������Ⱥ�к��ٵĸ������γ��˼��������ķ�֧��������ʾ������ʾ����ԭס���Ǵ�����׳����Ⱥ��̨��ԭס���и�������ġ����⣬����ֵĵ���Ⱥλ�ڶ����ķ�֧����Ȼ��¡�˵ĵ���Ⱥ�����ߵ��������γ���������С�ķ�֧��������ϵ͵IJ��ֵĺ�ɫ�ڵ㣩�������֧�ij߶���Խϴ���ռ�������������֮һ��
��O2a*�������У�����ķ�֧�ȴ�½��̨��IJ��ָ���˵�������DZȴ�½׳����Ⱥ��̨��ԭס������ϣ�������Դ�Դ�½׳����ϵ��Ⱥ�塣��STR������Щ��߶ȵķ�֧��ͨ���൱����ʱ����������γɵġ�
���ǹ�����Ⱥ���Ϊ��O1a*Լ3.6���꣬��O1a*�����к��Ϸ�֧�������Լ1.9���꣬��O2a*�������3.2���꣬��O2a*�����к��Ϸ�֧�������Լ2.6���꣨��2������ע�����Ƕ�O2a*�Ĺ�ֵ�ǻ��ڻ�����Ⱥ����˲�ͬ�����緶Χ��O2a*�����������������Ⱥͬ�����ٳ����ڻ������ڻ�����Ⱥ������������Ⱥ������ܿ��ܾ������ǽ��붫�ǵ�ʱ�䡣O1a*��O2a*������ܽӽ������ֱ���1.8�����2.6���ꡣ���ʱ������ĩ�α��ڸ߷������Ǻϣ���ʱ�й�����Ĵ�½���ں�ƽ�����϶��Ҷ��ִ��˽��붫����˵�ṩ��һ���ݾ���
��������Ǩ����붫�ǵ��ִ�����˵���ܴ���һ�����ϵ�ͨ·����һ�ֿ��ܵĽ��붫�ǵ�;���ǴӶ�����-���ǵ���������������鵽���Ϻ��й��ڵأ�ͼ1A�������Ǽٶ�O3����Ⱥ�Ǿ���������·���ɺ��غ���������Ⱥ�����ģ����ܻ�û���㹻��֤�ݣ�����������������ϸ�����Ⱥ���о������˿��ܴ������ߵ���������Ȼ����O3Ҳ������ͨ����ͬ��һ������·�߳��ֵģ���������O1a*��O2a*���ֵ�ʱ�䡣O3����Ⱥ����ͨ�����·ֲ�Ӱ���˺ܶ�������Ⱥ������O3�ں��ϵ�������Ⱥ�к������жϣ���׳���˵������״ν��붫��ʱ��û��O3�Ӻ��ϵ��������롣������Σ�����ȷ�������ض��߽��붫�ǵ�����Я����O2a*��һ������ϵĵ���Ⱥ������4���꣬����δ���������ݣ���O2a*����Ⱥ����Ҳ������ִﺣ�ϵ�����Ⱥ�����ϵ��ϵ�O2a*��֧�������ȷ��ԼΪ2.6����ǰ��������ݱȺ�������ϵ�������ʶ���ַ����Զ�������������Ǻ���ԭס�����������������ܻ��ں��ϵ����ָ���Ŀ�����ַ��
���ǵ�O1a*�������ͼ��ʾ̨��ڵ�Ⱥ��Ͻڵ���ӽ���������ģ������ڵ�����̨��ܽӽ��������Ⱥ����Դ�ء���ˣ�O1a*������Դ�ں��ϵ��Զ����������غ��ϣ�����ȷ������O1a*��֧��O2a*��֧�����ᡣO1a*ͬ��������5000��ǰ���й������غ�����ʯ��ʱ�����������ҵ������ǵ���ʯ��ʱ��ʼ��8000��ǰ��O1a*Ҳ��������ʯ���Ļ���ɢ�����ϵ��Ŀ��ܡ�O1a*�ں��ϵ������Լ1.8����ǰ���Ⱥ��ϵ���ʯ��ʱ����ַ��Լ6000��ǰ�������ϡ�һ�ֿ��ܵĽ�����������ʵ�ʼ�������ڵ��ﺣ�ϵ�֮ǰ��O1a*��Ⱥ�뿪O1a*����Ⱥ�壨������̨��ԭס���ȣ���ʱ�䡣���Ǽ����̨��ͺ���ԭס��O1a*֮��ķ���ʱ����2.2��95����������1.25-4.68�����ꡣ
��֮�����ǵķ��ֽ�ʾ�˺���ԭס����Դ��ĩ�α��ڽ��붫�ǵ�������������ʱ�����������������ԭס��û���ܵ����Ǵ�½����Ⱥ���·ֲ���Ӱ�죬����ԭס���Y-SNPͼ����ӽ�������Ŵ��ṹ��������Ϊ����ԭס����IJ����ܽ�ʾ�����˵���Դ�Լ����ǵ��Ŵ���ɫ������Ҳ���Ե���������Ⱥ�Ŵ��о���ģ�͡�
����֪�����������Ⱥ��һЩ�Ŵ�ѧ�о���˵�Ǻ����õġ����磬���Ӳ�֢���²�������Ȼ�ڸ�����Ⱥ�п��Ա��������ڷ��������⣬��Խϴ����Ⱥ�;�Զ����ʷʹ�ô��������ԭס��̫���ܷ����Ŵ�Ư�䣬ȴ���Է�չ�����Y-SNP��̬����Զ��Ը��ߵ�Y-STR��̬�ԡ�����ȷ�ź���ԭס��Ҳ���ű�����������Ⱥ���ߵij�Ⱦɫ������̬�ԣ����͵�������ƽ�⣨linkage disequilibrium�������ڼ����������о��п��������ɸ�������ƽ������Ĵ�������Խ������ˣ�������Ϊ�������Ŵ�ѧ�о��ж����Խ�����ԭס����Ϊ���Ǹ�����Ⱥ��ģ�͡�