![]()
|
|
|
| 从Y染色体和mtDNA角度分析朝鲜民族的父系母系起源问题 一,Y染色体类型――父系 朝鲜的Y染色体单倍体类型,主要是O2b和O3,其次为C3、D2、Q1、O1、O2*、N*、N1等类型,它们有着不同的起源方向。 东亚汉族、朝鲜、蒙古、满族、Evenk、日本六个人群的Y染色体单倍体类型树(图)
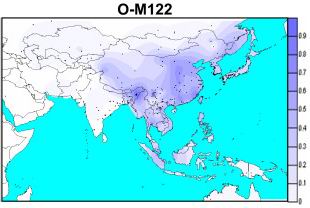
主干1――O2b,这个类型是朝鲜民族本民族的核心类型,在朝鲜族约占37.3%。这个人群在O2(P31)两个主要支系之一,大致是10~15KY前从东南亚地区北上,并和O2a人群分化,最终扩散到朝鲜半岛和日本列岛。 在朝鲜以外的地区,O2b主要分布在日本(31.9%)、越南(4.7%)、满族(3.8%)等民族中,可见O2b主要分布于朝鲜半岛和日本地区。 但日本人主要是O2b1,和朝鲜人的类型并不一致。目前公开发表的数据中,尚没有在汉族中发现O2b的报道。 O2b的近亲,是O2a,和原始型O2*,O2a主要分别中国南部和东南亚的很多苗瑶语(Hmong-Mien)、壮侗语(Daic)民族中,很多民族超过60%,在中国北部的中国鄂温克族、赫哲族中也有低频率出现,O2a在北方汉族出现频率小于3%,为华南人群向北扩散的结果。O2*原始型,则广泛分布在东亚地区,尤其汉族、朝鲜族、日本等人群中都有分布。 主干2――O3,O3在现在朝鲜民族中的出现频率高于40%,是朝鲜人群中出现频率最高的类型。但其起源却不是朝鲜本族的。 O3是东亚最主要的类型,分布也最广,从乌拉尔山到日本,从勘察加半岛到新西兰的毛利人中,都有O3的分布,最高频率出现在印度那加邦、中国云南地区和北部汉族人群中。 O3的主要类型是O3a5―M134(旧表示O3e),和O3a―LINE1(旧O3c+O3d),前者是汉藏语(Sino-Tibeto)民族标志性基因,后者在苗瑶语民族中达到最高频率。其他民族都是扩散的结果。 朝鲜人中的M134和LINE1的比例大体相当,由于LINE1目前看可能是东夷部族的类型,所以以此来看,现代朝鲜人中的O3应该是主要来自中国西北部的夏部落,少数来自华北东部的东夷部落。 O3在世界范围内的分布,华北、云南的扩散中心(图)
其他部分3――D2、C3、Q1、O1、N* 其中,D2只分布朝鲜半岛和日本地区,在日本Ainu人中达到最高,为87%,其次冲绳人,55%,日本本民族,为29.5%,在朝鲜人中,不同的取样有差异,但一般为3~6%,D类型是非常早达到的东亚的古老居民的后代,在东亚的分布非常广,汉族中的D大概为1%,主要是D3和D1,与朝鲜不同。 C3(M217),C在东亚分布很广,其中又以C3分布最广,C3可能最早分布在华北地区,O人群北上后,其才扩散到了东北,其后才扩散到蒙古高原和西伯利亚。在朝鲜人中比例为9.3%,与南部汉族相当,但高过北部汉族的(4.5%)、日本人(3.3%),两种可能,一是古代朝鲜有来自华南地区的影响,二是可能受到了东北地区一些民族的影响。如,俄国的Evenk人中C3为68.4%。但朝鲜族和西伯利亚的各民族没有关系,因为朝鲜人中完全没有C3c,,而西伯利亚的C3却基本是C3c,如Evenk为54.7%,两者不存在共同族源。 通古斯人标志的C3c的分布,在朝鲜人中不存在(图)
Q1(M120)是典型的华北汉族和一部分藏缅语人群的标志性基因,在各地汉族中出现频率在4%~10%之间,在境外只在越南人、中亚东干人和朝鲜人中发现过。在朝鲜人中,Q1出现频率1.3%。 O1(M119)是典型的百越基因,历史上百越可能是一个很大的民族,并且曾经拥有过很发达的文明,后期,汉语部落向东南扩展,百越开始朝不同方向迁徙,并给北亚、东北、东南亚各地带去了O1类型,在朝鲜人中,O1出现频率比较低,为2.1%。 N*和O是近亲类型,其发源地也是中国西南部地区,N的三个主要类型为N1、N2、N3,N1主要是东北亚地区(尤其是汉族和朝鲜族),N2为中亚和蒙古,N3为西伯利亚和通古斯各族,N3和N2在欧洲北部也有分布。朝鲜族中的N全部为N1,和北部汉族最接近。在西伯利亚各族中,N3的比例非常高,如Yakut人超过70%,而朝鲜人完全没有N3分布,两者没有族源关系。 综合看父系,可以分三部分,一、朝鲜民族具有本民族特点为O2b,占40%弱,来自1.5~1.0万前从华南北上居民的后裔,他们可能是朝鲜人语言的来源。二、来自华北地区人群的O3a5、O3*、Q1、N1占40%强,其进入朝鲜半岛的时间比较晚,且O3a5(M134比例)明显低于汉藏语民族,显示历史上曾经有来自华北东部的苗瑶语LINE1人群融入朝鲜半岛,且是很多人群分批进入的,所以对朝鲜民族的语言没有产生严重冲击。三、来自东北地区(或者华南)的C3和来自日本列岛的D2,占了10%。四、东亚人群共有O*、N*、O2*,以及后期来自华南的部分(O1、O2a),还有一部分来自西亚(R*),这些低频类型总计占了10%。 汉、朝鲜、满、蒙古、日、艾温基人(通古斯)六个人群的Y染色体单倍体类型比较(图)
总体看,朝鲜民族的Y染色体具有非常鲜明南部起源特征。 二、线粒体mtDNA――母系 东亚人群的母系可以划分为M系和N系统、R系统(N的一个子系统)三个。 其中M系统到达东亚的时间最早,其路线是沿着海岸前进的。其次是N系统,其路线可能是从印度出发,是人类第二大迁徙的结果。R系统是从中亚开始扩散的,代表了人类第三次大迁徙的结果。 东亚汉族、朝鲜、蒙古、满族、Evenk、日本六个人群的mtDNA类型树(图)
先行者――M M系类型主要包括M、D、C、Z、G,在全球范围内来看,M系最高频率出现分布在印度和西伯利亚。 在东北亚地区来说,一般规律是越向北,M系统所占比例越高,越向南频率越低,在五个民族中M(M、D、C、Z、G)出现频率分别是满族(80.9%)、日本人(70.8%)、朝鲜族(68.7%)、汉族(58.8%)、蒙古族(52.5%)。可见,M是东亚人群非常标志性的mtDNA类型,在所有人群中都超过了50%,满族更是高达80.9%。 M系中,最值得一提的C、D两个,C是典型的西伯利亚类型,在5个人群,中只有满族(18.4%)和蒙古族(12.5%)有这非常高的C的出现频率,表明这两个民族于西伯利亚人群有着非常紧密的联系。其他人群出现频率都小于3%。 D类型则更为奇特,它是东亚和美洲印第安人人非常多见的类型,在亚洲,D在西伯利亚和东南亚出现频率都很高,但恰恰在两者的中间――华北地区,出现频率却比较低。体现了人类第二次和第三次大迁徙对古代华北居民的母系结构产生过很大影响。 规模比较小的迁徙――N(不含R) N系统类型主要包括N、A、I、W、X、Y等类型,代表了人类在亚欧大陆第二次大迁徙的影响。 这次大迁徙对东亚人群的母系结构冲击要小很多,在五个人群中分别达到了蒙古(20.6%)、朝鲜(15.7%)、汉族(14.7%)、日本(10.1%)、满族(5.9%)。 在全世界范围内看,这次人口迁徙影响也不大,其出现频率最高为北美地区,如北美印第安人很多人群A的出现频率超过80%。 在上述东亚五个人群中,N主要出现在蒙古(12.5%)、汉族(6.6%)、朝鲜(5.1%),对日本和满族几乎没有影响,A主要出现在日本(10.1%)、朝鲜(7.4%)、蒙古(6.9%)、汉族(6.4%),满族中比较少见。 同时要说的是,A在汉族中的分布很不均匀,有的地区超过15%,有的地区则只有3%。 来自中亚的扩散――R(不讨论欧洲的HV、H、V、J、T、K、U) R类型迁徙主要影响了大陆西部的欧洲,但不是说对东亚就没有影响,尤其是华北和华南。 R的主要类型有R、F、B等等,在五个人群的出现频率为汉族(25.2%)、蒙古(20.6%)、日本(191%)、朝鲜(15.7%)、满族(11%)。 B类型广泛分布在亚洲,日本(11.3%)、汉族(11.1%)、朝鲜(10.6%)、满族(8.8%)、蒙古(6.9%),B类型在南美洲也有较广的分布,在南部汉族中,B的比例远超过北部汉族。 F主要出现在蒙古(12.5%)、汉族(12.1%),其他民族比较少见。F在全球范围内主要分布在乌拉尔山两侧、中亚、蒙古地区、东亚地区,在华南地区出现频率也非常高。 另外,欧洲居民比较常见的HV、H、V等在蒙古(6.3%)、满族(2.2%)、汉族(1.3%)也有部分的零星分布,显示了古代欧洲居民也曾有部分来到过东亚。 汉、朝鲜、满族、蒙古、日本五个人群mtDNA类型比较(图)
综合看母系mtDNA,朝鲜族M系占有十分明显的优势,从东亚地区M系北高南低的特点看,其起源地应当比汉族更靠北一些。 结论 朝鲜人说的“南男北女”从Y染色体和mtDNA上看,还是有一定道理。其父系明显是南部起源的人群占了主体,但其母系又不同于其南方的华北、华南人群。 |